
[doc] 细胞因子与发热机制研究进展
14页
1、细胞因子与发热机制研究进展?1156?细胞因子与发热机制研究进展*谢新华,综述董军审校暨南大学医学院病理生理教研室(广州510632);赣南医学院生理教研室(江西赣州341000)细胞因子是由免疫细胞和部分非免疫细胞合成和释放,具有细胞间信息传递,免疫调节作用的一大类小分子蛋白质或多肽.其中部分细胞因子与发热反应关系密切,在发热中起重要作用:有些促炎细胞因子(proinflanunatorycytoldnes)如几一1,IL一6,TNFCt等是重要的内生致热原,具有明显的致热作用;而另一些抗炎细胞因子(antiinflammatorycytokines)如IIJ一10,ILl-a具有解热作用.随着基因敲除技术,电生理技术等的发展和应用,细胞因子在发热中作用的研究取得了一些进展.1与发热相关的细胞因子1.1IL一1家族和几一1J3白介素一1家族包括几一1B,IIJ一1a,IL一1受体内源性拮抗蛋白(ILl-a).它们作用于相同的受体:白介素一1受体(几一1R).IL一1R分为工型(IL一1R工)和型(IL一1R),IL一1R工属于Toll样受体家族成员,几一1与IL一1RI结合后,通过T
2、0样受体信号转导途径促进细胞因子(几一1J3,IL一1a,IL一6,TNFCt等)的表达_lJ.IL一1R与IL一1也具有高亲和力,但与之结合后不能激活后续的信号转导途径.实验性小鼠,家兔,人静脉注射LPS均可引起发热,此时血浆和脑脊液几一1J3水平升高;LPS能上调IL一1J3,IL一1a,IL一1RI的表达,而通过抑制IL一1RI1mRNA的转录下调IL一1RI1表达.Kozak等利用IL一1B基因敲除(knockout,KO)技术发现,小剂量LPS(100ks,ip)诱导的发热中,KO小鼠与野生型(wildtype,WT)小鼠相比仅表现为小幅度抑制;而大剂量LPS(如2.5rag/kg,ip)诱导的发热中,KO小鼠24h内的发热峰值明显减小.这些实验证据均说明IIJ一1J3是一种重要的致热性细胞因子.1.2IIJ一1受体拮抗蛋白(interleuldn一1receptorantagonist,IL一1l-a)IL一1l-a与IIJ一1RI也具有较高的亲和力,但结合后不能激活后续的信号转导途径,是IL一1p的竞争性抑制剂.Cartmell等_2j在实验中预先给予IIJl-a或使IL
3、l-a在体内过度表达均可减轻IL一1J3引起的发热,延迟发热时间并提高对IIJ一1J3,IL一1a的耐受性;而皮下注射LPS引起局部IIJ一1J3,IIJ一1a浓度升高,2h后IIJ一1l-a水平升高,且IL一1m的浓度与发热程度呈负相关.Xin等在豚鼠视前下丘脑脑片实验中发现,人重组IIJ一1J3能减少热敏神经元的放电频*国家中医药管理局基金资助项目(编号:2003LHR13)2广东省中医药管理局基金资助项目(编号:1040074-);/x通讯作者率,增加冷敏神经元的放电频率,而几一1l-a能抑制IIJ一1J3的上述作用.由此看来,IIJ一1l-a是重要的内源性IL一1拮抗剂,具有限制发热的作用.1.3肿瘤坏死因子一ct(TNFa)TNFCt以三聚体形式存在于血液,并以三聚体与其受体I型(p55)和型(p75)结合.实验中给大鼠静脉注射一a可以诱导发热,且血浆IIJ一1,几一6浓度升高,说明TNFa有致热作用,其机制之一可能是通过诱导IIJ一1,IIJ一6的合成.Leon等用TNF受体基因敲除技术对小剂量LPS和松节油引起的发热进行研究,发现TNFKO小鼠和WT小鼠体温变化均无明显差
4、异,提示TNF在小剂量LPS和松节油致发热中似乎不起作用.1.4白介素一10(一10)几一10是内源性抗炎细胞因子.Camnell等_3J发现给小鼠腹腔和脑室内注射鼠重组型IL一10能抑制革兰阳性和阴性细菌毒素所致发热,并伴有血浆IIJ一6水平降低,而中和IIJ一10则使发热时程延长,水平升高.用基因敲除技术进一步表明,不论是大剂量还是小剂量LPS所致发热,IIJ一10KO小鼠的发热峰值和时程均高于WT小鼠,且血浆中几一6水平也高于WT小鼠;但在松节油引起的发热,IL一10KO小鼠与WT小鼠体温变化无明显差别,提示IIJ一10可能不参与松节油性发热.几一10限热作用的机制可能为IL一10与几一10受体结合后通过JakStat信号转导途径,抑制促炎性细胞因子IIJ一6,IIJ一1,TNFa等的合成来实现.1.5白介素一6(IIJ一6)传统实验方法得到的大量实验结果表明,IL一6是重要的致热性细胞因子,随着基因敲除技术的应用,进一步得到证实.Kozak等发现用小剂量LPS可引起wT小鼠中等程度的发热,但未能引起IIJ一6KO小鼠出现发热;大剂量LPS使两种小鼠出现相似的发热及炎症反应:说明
《[doc] 细胞因子与发热机制研究进展》由会员re****.1分享,可在线阅读,更多相关《[doc] 细胞因子与发热机制研究进展》请在金锄头文库上搜索。

外科手术部位预防与控制试题35题

企业年会主持稿5篇

Me小学英语作文范文

日常生活安全防范小常识

物业工程部考试题集锦

卫生局医政科工作职责

2023年暑假学习计划范本(八篇).doc

贵州省六盘水市第十三中学九年级政治全册第12课找准自己的位置导学案无答案人民版
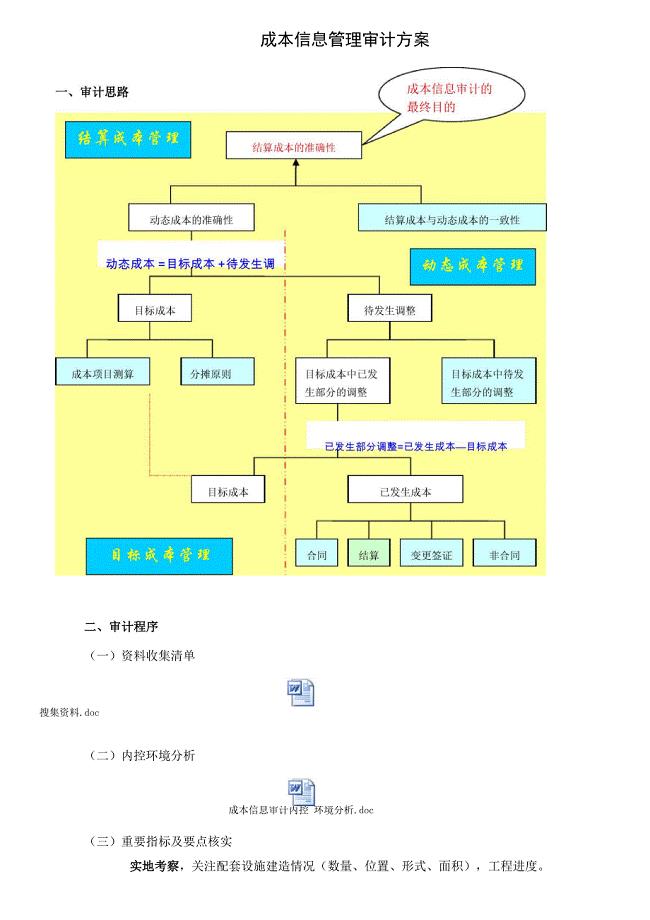
房地产公司成本审计方案

学困生帮扶年度工作计划标准范文(6篇).doc

工程装修施工合同(通用6篇)

小学三年级数学《毫米、分米的认识》教案范本三篇

lwpbA002LED点阵显示屏系统设计及PC机的串行通信

历代高僧放生诗略解

小学二年级科学上册期末测试卷

化工原理(第三版_陈敏恒)下册课后答案

教师下半年目标计划范本(五篇).doc

全息存储技术详解

2022年上海市建筑三类人员项目负责人【安全员B证】考试历年真题汇总含答案参考87

2023有关经典辞职报告锦集6篇
 廉租房协议书官方
廉租房协议书官方
2023-09-27 9页
学生成绩管理系统详细操作过程汇编
2023-09-20 17页
以主题式综合实践为载体实现多学科统整的实践研究
2023-04-20 14页
混凝土课程设计[共5篇]
2024-02-02 13页
1.1.1算法的概念
2022-09-02 7页
素食餐饮店的创业计划书
2023-11-07 10页
注册监理工程继续教育考试(房屋建筑)
2022-09-30 24页
装饰装修工程质量验收用表
2023-12-17 38页
《骆驼祥子》读后感1500字左右(《骆驼祥子》读后感怎么写)
2023-12-18 4页
房地产公司成本审计方案
2023-09-28 3页